Oxygen and biomass: an overlooked constraint
Why internal oxygen supply may matter more than we think in crop productivity
The simple story is incomplete
Plants are usually described as oxygen producers.
That is true at the scale of the whole plant and the biosphere. Through photosynthesis, plants use light energy to fix carbon dioxide and release oxygen. Without this net gain in carbon, there is no biomass accumulation. In that sense, productive crops are clearly net oxygen producers.
But this familiar story hides an important biological detail.
Every living plant cell also uses oxygen.
Plant cells require oxygen for mitochondrial respiration. Respiration is the process that converts stored carbon compounds into usable energy, reducing power and metabolic intermediates. These are needed for growth, maintenance, nutrient uptake, biosynthesis, transport, stress responses and developmental processes.
So the real question is not simply whether a plant produces oxygen.
The more interesting question is whether the right tissues receive enough oxygen, at the right time, to support the metabolic work required for growth and biomass formation.
Plants do not solve oxygen supply like animals do
Animals solve the problem of internal oxygen supply through circulation.
Oxygen enters through specialised exchange surfaces, binds to transport molecules, and is rapidly distributed through a circulatory system. This allows metabolically active tissues deep inside the body to maintain high rates of respiration.
Plants solve the problem differently.
Plants do not pump oxygen to every internal cell. They rely on diffusion, internal air spaces, tissue porosity, developmental structure and metabolic adjustment. This is a very different design philosophy.
Some plant tissues form internal air spaces that improve gas movement. Some develop porous structures or lenticels that allow exchange with the atmosphere. Roots under waterlogged conditions may form aerenchyma, creating channels through which oxygen can move. Some tissues reduce growth or lower metabolic activity when oxygen supply becomes limiting.
This means plant oxygen supply is not only a question of photosynthesis. It is also a question of structure, distance, tissue density, developmental stage and respiratory demand.
Oxygen production is not the same as oxygen availability
A crop canopy may be photosynthetically active and still contain tissues where oxygen availability is limited.
This distinction is important.
Leaves exposed to light may release oxygen during photosynthesis. But growing meristems, developing stems, storage tissues, roots, seeds, tubers and dense internal tissues may not have direct access to that oxygen. They depend on diffusion from surrounding air spaces, external tissues or the atmosphere.
In bulky organs and dense tissues, diffusion distances increase. At the same time, respiratory demand can be high. Cells are actively dividing, expanding, importing sugars, synthesising cell walls, accumulating storage products or maintaining ion gradients.
This creates a possible mismatch:
The plant may be a net oxygen producer, while specific internal tissues experience local oxygen limitation.
That local limitation may not always be severe enough to cause obvious injury. It may simply reduce metabolic efficiency, slow growth, alter carbon use or change developmental priorities.
For biomass productivity, those subtle effects may matter.
Biomass is not just carbon capture
Crop productivity is often discussed as if the central limitation is carbon fixation.
That is understandable. Photosynthesis is the entry point for carbon into the plant. Improved light interception, photosynthetic rate and canopy duration are all important.
But biomass formation is not completed at carbon fixation.
Fixed carbon must be transported, unloaded, metabolised and converted into structure or storage. This requires energy. It requires respiration. It requires oxygen.
Cellulose synthesis, lignification, sucrose storage, protein turnover, nutrient uptake, phloem loading and unloading, membrane transport and growth all depend on active metabolism.
A plant does not become productive simply because carbon enters the system. It becomes productive when carbon is successfully converted into useful biomass.
That conversion step may be where overlooked constraints sit.
The oxygen ledger
One useful way to think about plant productivity is as an oxygen ledger.
On one side of the ledger, photosynthesis produces oxygen as carbon dioxide is fixed.
On the other side, respiration consumes oxygen as carbon compounds are oxidised to release energy and support metabolism.
The plant gains biomass when carbon fixation exceeds carbon loss through respiration and other processes. But this net gain does not mean that oxygen supply is irrelevant. It only means that, at the whole-plant level, carbon gain is positive.
Within the plant, the situation is more complex.
Some tissues are sources of oxygen and carbon. Other tissues are sinks for carbon and oxygen. Some tissues are photosynthetic. Others are not. Some are well aerated. Others may be dense, wet, buried, enclosed or metabolically intense.
The oxygen ledger is therefore not evenly distributed across the plant.
A crop can have a positive whole-plant carbon balance while still facing local oxygen bottlenecks in tissues that are crucial for yield.
Sink strength may depend on oxygen supply
In crop physiology, we often discuss the importance of sink strength.
A strong sink is a tissue or organ that attracts, imports and uses assimilated carbon. Examples include developing stems, grains, fruits, roots, tubers and storage organs.
Sink strength is often described in terms of carbon demand, enzyme activity, transport capacity, storage capacity or developmental priority. These are all important.
But sink activity also requires energy.
To import sugars, maintain gradients, synthesise polymers, build cell walls, accumulate storage compounds and sustain growth, sink tissues must respire. That means they need oxygen.
If oxygen supply is inadequate, sink metabolism may slow. Carbon may accumulate elsewhere. Growth may be reduced. Development may shift. The plant may appear to have a carbon allocation problem, when part of the issue is actually an energy and oxygen supply problem.
This raises an important possibility:
Some limits to biomass accumulation may not be caused by inadequate photosynthesis, but by the capacity of sink tissues to use carbon efficiently under their local oxygen environment.
Sugarcane as an example
Sugarcane is a useful crop for thinking about this problem.
It produces large amounts of biomass and stores high concentrations of sucrose in stem internodes. The crop is often discussed in terms of canopy photosynthesis, stalk growth, sucrose accumulation and biomass partitioning.
But the sugarcane stem is not a simple storage tube.
It is a complex, living structure. Internodes differ in age, water content, fibre composition, sucrose concentration, respiratory activity and developmental state. Younger internodes are actively growing and metabolically demanding. Older internodes may accumulate sucrose and become structurally more mature.
The question is whether oxygen supply inside these tissues is always adequate to support the metabolic processes required for maximum biomass and sucrose accumulation.
This is not usually the first question asked in sugarcane improvement or biomass research. We tend to ask whether the canopy captures enough light, whether the crop fixes enough carbon, whether there is enough genetic variation, whether sucrose storage is strong enough, or whether fibre composition can be improved.
Those questions remain valid.
But we may also need to ask:
- Are rapidly growing internodes sufficiently oxygenated?
- Does internal tissue structure influence respiratory efficiency?
- Does oxygen availability affect sucrose accumulation, fibre formation or internode maturation?
- Are there genotypic differences in tolerance of low internal oxygen?
- Could some high-biomass phenotypes be better at managing internal oxygen supply?
These questions may open a different way of thinking about biomass constraints.
Poor oxygen supply does not always mean flooding
When plant scientists hear “low oxygen”, they often think of waterlogging or flooding.
That is understandable. Flooded soils limit gas exchange, reduce oxygen supply to roots and can cause major stress. Aerenchyma formation and anaerobic metabolism are well-known responses to these conditions.
But oxygen limitation does not have to be restricted to flooded roots.
Low oxygen can also be a local internal condition. It can occur where tissues are dense, diffusion distances are long, respiration rates are high, or gas spaces are limited. It may occur transiently during rapid growth, at night, under high temperature, or in tissues with intense metabolic demand.
This means oxygen limitation may be more common, more subtle and more developmentally specific than we usually assume.
The issue may not be dramatic oxygen starvation. It may be marginal oxygen limitation that quietly reduces metabolic performance.
For crop improvement, that distinction matters.
Biomass efficiency and oxygen use
A key unresolved question is whether biomass productivity differs among genotypes partly because they differ in oxygen use efficiency or low-oxygen tolerance.
Some plants may maintain growth better when oxygen supply is restricted. Some may have tissue structures that improve internal gas movement. Some may regulate respiration more effectively. Some may maintain sink activity under conditions where other genotypes slow down.
If this is true, then oxygen-related traits could contribute to biomass productivity.
This would not replace photosynthesis, canopy development, water use, nutrient uptake or carbon allocation as major areas of crop research. Rather, it would add another layer.
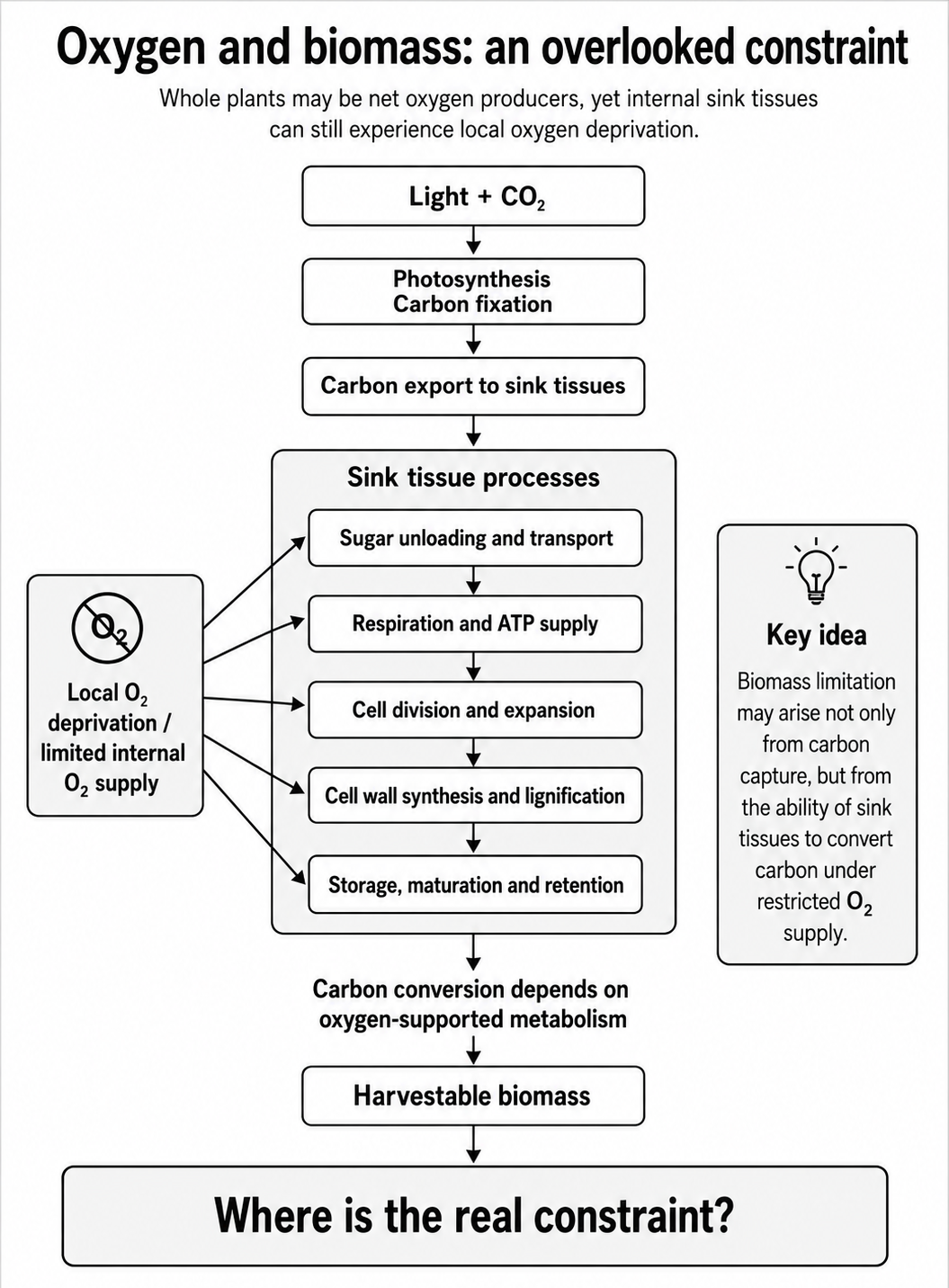
Biomass formation depends on an integrated system:
Oxygen may influence several steps in this chain, especially sink metabolism, growth and storage. As shown in Figure 1, oxygen may influence several sink-tissue processes rather than a single step in the biomass formation pathway.
Why this may be overlooked
Oxygen is easy to overlook because it appears obvious.
Plants produce oxygen. Therefore, we assume oxygen is not limiting.
But that assumption may be too simple.
Oxygen availability is spatially variable inside plants. It depends on tissue anatomy, developmental stage, water content, temperature, respiration rate and diffusion path length. It may change between day and night. It may differ between organs. It may differ between genotypes.
Another reason oxygen is overlooked is that it sits between disciplines.
It is not purely photosynthesis. It is not purely respiration. It is not only stress physiology. It is not only anatomy. It is not only crop modelling. It cuts across all of these.
That makes it easy to miss in R&D pipelines that are organised around familiar categories.
Implications for R&D strategy
If oxygen supply contributes to biomass limitation, then several practical implications follow.
First, biomass research should not focus only on carbon capture. It should also consider carbon conversion. The ability to use fixed carbon efficiently may be as important as the ability to capture it.
Second, sink tissues deserve more attention. In high-biomass crops, the tissues that build, store and retain carbon may be where important constraints are expressed.
Third, internal tissue structure may have hidden value. Traits such as porosity, vascular arrangement, cell size, water content, diffusion pathways and developmental anatomy may influence oxygen movement and respiratory performance.
Fourth, technology pipelines should begin with the constraint. If the real bottleneck is sink metabolism or internal oxygen supply, then adding more upstream discovery projects on carbon fixation may not deliver the expected productivity gains.
Fifth, realistic timelines matter. Discovery projects that target complex biomass traits must be honest about the distance between mechanism, trait identification, breeding, field validation and commercial delivery.
This is where strong R&D portfolio review becomes valuable. It helps ask whether projects are aligned with the actual productivity constraint, or whether they are simply technically interesting.
A working hypothesis
The central hypothesis can be stated simply:
In high-biomass crops, internal oxygen supply and respiratory efficiency may place overlooked limits on the conversion of fixed carbon into harvestable biomass.
This does not mean oxygen is always limiting. It does not mean photosynthesis is unimportant. It does not mean all biomass problems are oxygen problems.
It means that oxygen should be considered as part of the crop productivity system.
The more specific hypothesis is that some tissues, under some developmental conditions, may experience oxygen environments that constrain sink metabolism and biomass formation. Genotypes that manage this better may have an advantage in biomass productivity, especially under conditions of rapid growth, high temperature, dense tissue development or strong sink demand.
Questions worth asking
Several research questions follow from this view:
- How does oxygen concentration vary within stems, storage tissues, roots and developing organs during growth?
- Are there consistent differences among genotypes in internal oxygen status?
- Do high-biomass genotypes differ in tissue porosity, diffusion pathways or respiratory regulation?
- Does low internal oxygen affect sucrose accumulation, fibre synthesis or cell wall composition?
- Can oxygen-related traits be linked to biomass efficiency under field conditions?
- Are current crop models missing an important constraint by treating respiration too generally?
- Can breeding or biotechnology improve the capacity of sink tissues to function under marginal oxygen supply?
These are not trivial questions. They require careful measurement and good experimental design. But they may help explain why some productivity gains remain difficult to achieve despite major advances in photosynthesis, genomics and crop management.
Where is the real constraint?
In crop improvement, one of the most important strategic questions is:
Where is the real constraint?
If the constraint is light interception, then canopy architecture matters.
If the constraint is photosynthetic rate, then leaf-level carbon fixation matters.
If the constraint is water or nutrient supply, then root systems and agronomy matter.
If the constraint is sink capacity, then storage tissues, developmental regulation and carbon unloading matter.
But if the constraint is the ability of internal tissues to sustain the metabolic work of growth under limited oxygen supply, then we need to look in a different place.
We need to look inside the plant.
Not only at the genes that control growth, or the enzymes that process carbon, but at the physical and metabolic environment in which those processes occur.
Conclusion
Plants are net oxygen producers, but this does not make oxygen irrelevant to plant productivity.
Every living plant cell uses oxygen. Every growing sink requires respiration. Every storage process has an energy cost. Every dense tissue creates a physical environment in which gas movement may matter.
For high-biomass crops, the overlooked question is not whether the plant produces oxygen.
The question is whether oxygen reaches the right cells, in the right tissues, at the right time, and in sufficient amounts to support the conversion of carbon into biomass.
That may be one of the hidden constraints in crop productivity.
And if it is, then oxygen deserves a more central place in how we think about biomass, sink strength and the next generation of crop improvement strategies.